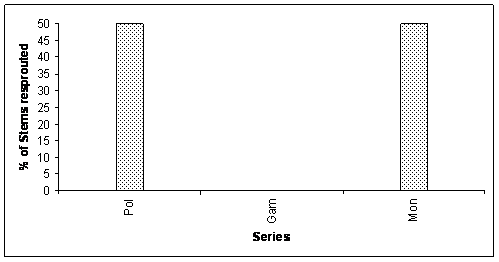|
Ethnobotanical Leaflets 13: 59-64. 2009.
Resprouting in Tropical Rainforest of Highwavy Mountains, Western Ghats, Tamil Nadu, India
Jegan, G. and Muthuchelian, K.
Centre for Biodiversity and Forest Studies, Department of Bioenergy, School of Energy, Environmental and Natural Resources, Madurai Kamaraj University, Madurai- 625 021,Tamil Nadu, India
Issued 04 January 2009
ABSTRACT Six hectares of evergreen forest of Highwavy Mountains, Western Ghats, Theni District, Tamil Nadu, India, was censused for the damaged trees (≥ 30cm girth at breast height). Among 57 damaged trees 28.07% was uprooted and 71.93% was standing broken stems. Among standing broken stems only 39.02% was resprouted. The percentage of resprouting of our study site was slightly higher than other forests. The resprouting and the production of multishoots from the stumps were mainly seen in the pioneer species and resprouting was rare in the climax species. This was contrast to other forests. Key words: resprouting, stumps, tree damage, rainforest, Western Ghats.
INTRODUCTION The death of the whole tree affects nutrient cycling, regeneration, and species richness in the particular area. Tree falls are determined by local climatic factors, physical characteristics of the substrate, and biological attributes of trees (Brokaw 1982, Putz & Milton 1982, Putz et al. 1983, Denslow 1987, Putz & Brokaw 1989). When a whole tree falls to the forest floor, it; cause pulses of organic material and nutrients that can subsequently become available to terrestrially rooted plants (Denslow 1987); increase the biomass of the forest floor, thus creating additional habitats for terrestrial organisms. Crush seedlings, saplings and understory plants (Aide 1987; Gartner 1989; Kinsman 1990); and affect microclimate of the ensuing gap that may daunt or smooth the progress of seed germination of some species (Putz & Milton 1982, Brandani et al. 1988, Swaine et al. 1990). While many damaged trees die, some continue to live by producing new shoots from above or below ground parts. Most of the research has centered on regeneration from seeds, seedlings, or clonal growth (Clark & Clark 1989, Eriksson & Ehrlen 1992). Resprouting from standing broken stems might replace the lost canopy and affect the form and duration of gap regeneration faster than regeneration from seedlings. The ability to resprout might allow a species to maintain its frequency in the population (Knight 1975, Putz & Brokaw 1989, Whigham et al. 1991). The rates and frequency of tree damage expected to be greater in higher elevation forests because of steeper slopes, less stable soil and exposure to more wind. Plant adaptations to these environmental characteristics might also be expected in wet evergreen forest. In this paper, we describe the damage of the trees and report the frequency of resprouting of snapped trees of evergreen forest of Highwavy Mountains, Western Ghats, Theni District, Tamil Nadu, India.
METHODS Study Site Our study site is situated in the Pachakumachi hill (9° 35’ to 9° 45’ N latitude and 77º 15’ to 77º 27’E longitude) of Western Ghats, South India. The Pachakumachi hill is surrounded by Palani Hills in the North, Sethur and Sivagiri hills in the South, Cardamom hills and Kerala state in the West, the Varushanadu hills in the Northeast and Thekkadi hills in the Southwest. Vaigai and Surliyaru are the main rivers originating from Pachakumachi hill ranges.
Climate and Soil Climatological data of the study site are collected from Pachakumachi estate Climatological station. Pachakumachi hill receives 2726 mm rainfall annually. June is recorded as the hottest month with maximum temperature of 31°C and January is the coldest month with the minimum temperature of 17° C. Humidity is high (95%) during the months of June, July and August; and low humidity is noted only in the month of March (85%). Climatological data of Pachakumachi hill is given in Fig 1. Pachakumachi hill has red, sandy clayey and loamy type of soil. Physio- chemical characteristics of soil of the study site are given in table 1.
Vegetation In the 10,000 Acres of total area of Pachakumachi hill, 2,000 acres are under the cultivation of cash crops such as cardamom, coffee and tea. These plantations are intermingled with the patches of Evergreen forests. The altitude of the hill ranges from 600 m to 2,000 m. The vegetation ranges from scrub jungles in the foothill to evergreen and sholas at hill tops. Our study site is situated at an altitude of 1,700 m. Our study site is defined as the tropical evergreen forest.
Field Work Between June 2006 and February 2007, we censused the entire site (6 hectares) for snapped or fallen trees and resprouting of the standing broken stems during study period. We made frequent visits to the study site. The damaged trees with GBH ≥ 30 cm were noted. Tree damage was divided into three categories; a: uproot- fallen trees with exposed root balls; b: knockdown- a broken or uprooted, tree falling as a result of a neighboring tree hitting it; c: standing dead- tree dead, but stem not broken or uprooted;. During study period, all standing broken stems were checked for the evidence of resprouting of leaves anywhere on the stem. Standing broken stem with newly sprouted leaves were recorded as living ones and those with no sprouting of leaves were recorded as dead ones. The resprouted stumps were classified into two groups; by the formation of new shoots; 1: Multishooted stumps- The stump producing more than one new shoots; 2: Monoshooted stumps- The stump producing only one new shoots; for this the standing broken stumps with DBH ≥ 30 cm were tagged. The stumps which were produced the shoots more than two in numbers, also noted. The stumps with DBH ≥ 30 cm were noted.
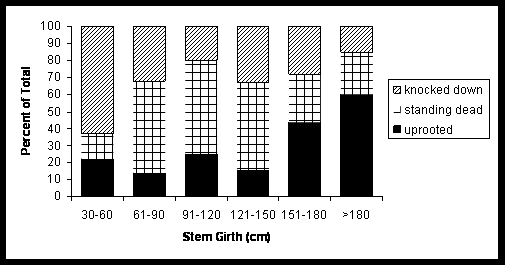
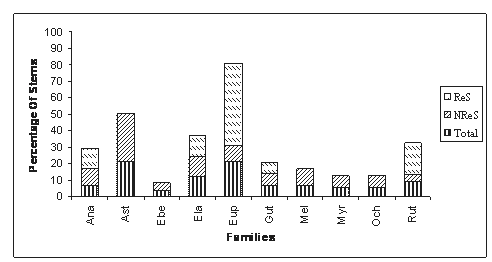
RESULTS In 6 ha study plot, 57 trees were severely damaged. The number of damaged trees per hectare was 9.5 trees. Among these, 28.07% were uprooted, 7.01% were standing dead, and remaining was knocked down (Fig 1.). All the trees that were damaged or died during our study period (57) were identified up to species level. They were belonging to ten families, represented by ten genera, and ten species. The percentage of damage was seen in both cases i.e., in climax and in pioneer species. The uprooting of whole trees mainly occurred in the climax species (20). All the members’ of pioneer species were standing broken stems. The standing dead trees were the climax species. Resprouting of damaged trees: The resprouting mechanism is mainly seen in the pioneer species. Among the climax species only 4 species were resprouted. The percent of resprouted and not resprouted standing stems was shown in the Fig 2. Other 5 species didn’t resprouted. They were considered as the standing dead trees. The resprouting mechanism is mainly seen in the pioneer species. Among the climax species only 4 species were resprouted. The percent of resprouted and not resprouted standing stems was shown in the Fig 2. Other 5 species didn’t resprouted. They were considered as the standing dead trees. Euphorbiaceae family showed 50% of resprouting among total percentage of resprouting. Resprouting percentage for other families were 6.25, 12.5, 12.5, and 18.75 of Guttiferae, Anacardiaceae, Elaeocarpaceae and Rutaceae respectively. Polypetalae and Monochlymadeae shared the percentage of resprouting (Fig 3). The production of multi shoots was seen only in the pioneer species. It was not seen in the climax species. Among 8 of the resprouting pioneer species, 2 only producing more than 2 new shoots from the standing broken stem.
DISCUSSION The rate of resprouting in our study site was coinciding with other forests. There was a trend that, in the mature forests, gap-colonizing species suffer higher nonresprouting rates than other taxa. But our result was controversy to the above trend, i.e., in our study site the pioneer species showed the higher resprouting rate. Among the climax species only 4 species showed the resprouting. Likewise the resprouting, the production of multishoots was seen only in the gap-colonizing species. No other standing broken stems belonging to the climax species was producing the multishoots.
ACKNOWLEDGEMENTS We thank UGC for their financial support of this project. We also thank Selvam for his help during the field study. Our sincere thank to Tamil Nadu Forest Department for their permission.
REFERENCES Andrew N. Gray, S. J.Harold, Zald, Ruth A. Kern, and Malcolm North 2005. Stand Conditions Associated with Tree Regeneration in Sierran Mixed-Conifer Forests Forest Science 51(3):198 –210. Brokaw, N.V.L. and R.T. Busing, (2000). Niche versus chance and tree diversity in forest gaps. Trends in Ecology and Evolution 15: 183-188. Burslem, D.F.R.P. and T.C. Whitmore, 1999. Species diversity, susceptibility to disturbance and tree population dynamics in tropical rain forest. Journal of Vegetation Science 10: 767-776. Capellotto Costa, F.R., C. Senna, and E.M. Nakkazono. 2002. Effects of selective logging on populations of two tropical understory herbs in an Amazonian forest. Biotropica 34: 289-296. Canham, C.D.1989Different responses to gaps among shade tolerant tree species. Ecology, v. 70, 548-550. Denslow, J. S. 1980 Gap partitioning among tropical rainforest trees. Biotropica, v. 12 (Supplement), 47-55. Emborg, J. 1998: Understorey light conditions and regeneration with respect to the structural dynamics of a near-natural temperate deciduous forest in Denmark. Forest Ecology and Management. 106:83-95. Foster, M. S., J. Terborgh 1998. Impact of a rare storm event on an Amazonian forest Biotropica 30(3): 470-474. Goldblum, D. 1997. The effects of tree fall gaps on understory vegetation in New York State. J. Veg. Sci. 8: 125–132. Madsen, P. 1995b. Effects of seedbed type on wintering of beech nuts (Beech) and deer impact on sprouting seedlings in natural regeneration. Forest Ecology and Management. 73:37-43. Magnusson, W. E. et al.1999 Logging activity and tree regeneration in an Amazonian forest. Forest Ecology and Management, v. 113, 67-74. Malanson, G P (1996) Effects of Dispersal and Mortality on Diversity in a Forest Stand Model, Ecol. Model., 87: 103–110. Moore, M.R., and Vankat, J.L. 1986. Responses of the herb layer to the gap dynamics of mature beech–maple forest. Am. Midl. Nat. 115: 336–347. Nadkarni, N. M., J. Mateison, and Rodrigo Solano. 1995. Tree damage and annual mortality in a Motane Forest in Monteverde, Costa Rica. Biotropica 27(4): 441-447. North, M., J. Chen, B. Oakley, B. Song, M. Rudnicki, A. Gray, and J.Innes. 2004. Forest stand structure and pattern of old-growth western hemlock/Douglas-fir and mixed conifer forests. For. Sci. 50:299 –311. North, M., M. Hurteau, R. Fiegener, and M. Barbour. 2005. Influence of fire and el nin˜o on tree recruitment varies by species in Sierran mixed conifer. For. Sci. 51:185–195. Peterson C. J., Carson W. P., McCarthy B. C. & Pickett S. T. A. 1990 Microsite variation and soil dynamics within newly created tree fall pits and mounds. Oikos 58: 39–46. Sheil, D., D.F.R.P. Burslem, and D. Alder, 1995. The interpretation and misinterpretation of mortality-rate measures. Journal of Ecology 83: 331-333. Sheil, D. 2001. Long-term observations of rain forest succession, tree diversity and responses to disturbance. Plant Ecology 155: 183-199. Valverde, T., and Silvertown, J. 1998. Variation in the demography of a woodland understorey herb (Primula vulgaris) along the forest regeneration cycle: projection matrix analysis. J. Ecol. 86: 545–562. Van der Meer, P.J. and Bongers, F. 1996. Formation and closure of canopy gap in the rain forest at Nouragues, French Guiana. Vegetatio 126: 167-179. Van Pelt, R, and J.F. Franklin. 2000. Influence of canopy structure on the understory environment in tall, old-growth, conifer forests. Can. J. For. Res. 30:1231–1245. Wu, X. McCormick, J.F. and Busing, R.T. (1999) Growth patterns of Picea rubens prior to canopy recruitment. Plant Ecology, 140, 245–253. Welden, C.W. Hewett, S.W. Hubbell S.P., and Foster, R.B. (1991) Sapling survival, growth and recruitment: relationship to canopy height in a neotropical forest. Ecology, 72, 35-50. Zang, R.G. and Wang, B.S. (2002) Study on canopy disturbance regime and mechanism of tree species diversity maintenance in the lower subtropical evergreen broad-leaved forest, South China. Plant Biosystems, 136 (2), 241-250.
Table 1: Physio- chemical characteristics of soil of the study site.
Figure 1: Size class distribution (percent of total stems) of the damager stems.
Figure 2: The composition of damaged trees by family. The percent of total trees damaged, not resprouted and resprouted. (ReS- Resprouted, NReS- Not resprouted).
|